

 English
EnglishRESUMEN
El género Pyricularia es conocido por atacar principalmente gramíneas. Este hon- go presenta una amplia gama de hospederos entre los cuales se destacan el arroz y el trigo, así como otras gramíneas cultivadas, nativas y malezas. Hasta el momento poco se conoce sobre la importancia del inóculo procedente de las malezas para el cultivo del trigo. El objetivo de este estudio fue identificar los posibles hospede- ros alternativos de Magnaporthe oryzae, en las principales zonas de producción de trigo en Paraguay. Para ello, muestras de malezas, con síntomas característicos del ataque del hongo, fueron recolectadas en los departamentos de Itapúa, Alto Paraná y Canindeyú. Estas muestras fueron sembradas sobre medio de cultivo PDA, e incubadas durante 5 días a 25 ºC en el laboratorio de Fitopatología del Centro de Investigación Hernando Bertoni, Caacupé. Posteriormente se realizó la identifi- cación del hongo mediante observación al estereoscopio y microscopio. Con los aislados obtenidos, se realizaron pruebas de patogenicidad sobre trigo. Se constató la presencia del hongo en: Bromus catharticus, Brachiaria sp., Chloris gayana, Cype- rus diffusus, Digitaria horizontalis, Lolium multiflorum, Cenchrus echinatus, Avena strigosa, Digitaria insularis, Rynchelyntrum roseum y Eleusine indica. Se observó pa- togenicidad de los aislados de B. catharticus, L. multiflorum, C. echinatus y E. indica.
Palabras clave: Magnaporthe oryzae, trigo, hospederos alternativos
INTRODUCCIÓN
El género Pyricularia es conocido por atacar principalmente gramíneas, siendo el arroz el principal cultivo afectado por este patógeno a nivel mundial. Este hongo presenta una amplia gama de hospederos entre los cuales se destacan numerosas gramíneas cultivadas, nativas y malezas.
De acuerdo con Reis et al. (2005), el hongo se perpetúa de un año a otro en los re- siduos de cultivo, semillas y plantas espontáneas que pueden actuar como fuente de inóculo; según Lima (2004) son hospederos alternativos la cebada, milleto, maíz, triticale, centeno y acevén. Existen reportes del ataque de este hongo en Brachia- ria brizantha y Brachiaria extensa (Gutierrez et al. 2000; Verzignassi et al. 2012). Prabhu et al. (1992) y Urashima et al. (1993) reportan varias malezas de la fami- lia Gramineae: Cencrhus echinatus, Eleusine indica, Digitaria sanguinalis, Brachiaria plantaginea, Echinocloa crusgalli, Pennisetum setosum, Hyparrhenia ruffa y Rhynche- lytrum roseum, como posibles fuentes de inóculo del hongo. En el 2006, Mehta et al., habían agregado a la avena negra (Avena strigosa) a la lista de fuentes probables de infección primaria y Marangoni et al (2013) agregaron a la avena blanca (Avena sativa) como especie susceptible al brusone de trigo; así también Chávez y Kohli
(2015), confirmaron la patogenicidad de aislados de Bromus catharticus, Digitaria horizonthalis y Lolium multiflorum sobre plántulas de trigo, y reportaron la presen- cia del hongo en Cyperus diffussus y Chloris gayana. Con el fin de aclarar el papel que cumplen las malezas como fuente de inóculo de Pyricularia para el trigo, este estudio buscó identificar los posibles hospederos alternativos de Pyricularia sp. en las principales zonas de producción de trigo de Paraguay, y determinar si los aisla- dos de los mismos son capaces de infectar al trigo.
MATERIALES Y MÉTODOS
Durante los años 2013 a 2016, muestras de hojas e inflorescencias de malezas de las familias Poaceae y Cyperaceae, que presentaban síntomas característicos del ataque de Pyricularia, fueron recolectadas de las zonas aledañas a parcelas de tri- go, en los departamentos de Itapúa, Alto Paraná y Canindeyú.; así también se tomaron algunas muestras en el Centro de Investigación Hernando Bertoni en el departamento de Cordillera. Las hojas que presentaban manchas elípticas, con borde marrón rojizo u oscuro y centro más claro fueron tomadas para su análisis en el laboratorio, (Figura 1A) (Reis et al. 2005), así también las inflorescencias con lesiones necróticas cubiertas por crecimiento micelial grisacéo (Figura 1B).

Síntomas característicos del ataque de Pyricularia en hojas (A) e inflorescencias (B). Capitán Miranda, Paraguay, 2014.
En el cuadro 1 se detalla la lista de especies recolectadas en los diferentes departamentos.
Tabla 1. Lista de especies de malezas recolectadas en los departamentos de Itapúa, Alto Paraná, Canindeyú y Cordillera durante los años 2013 a 2016. 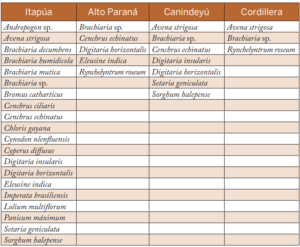
Las muestras fueron procesadas en el Laboratorio de Fitopatología del Centro de Investigación Hernando Bertoni de la siguiente manera: pequeñas porciones de la parte afectada fueron desinfectadas mediante inmersión en soluciones de alcohol al 70% durante 30 segundos, luego en hipoclorito de sodio (55 g Cl/l): agua (1:3), por un minuto, posteriormente lavados tres veces con agua destilada esterilizada y secados sobre papel absorbente. Una vez secos se procedió a sembrar los mismos en placas de Petri con medio de cultivo PDA (Papa, dextrosa, agar); e incubados a 25°C durante 5 días. Transcurrido el periodo de incubación, se procedió a la identificación mediante observación al microscopio utilizando la clave de Barnett y Hunter (1998).
Las colonias identificadas como Pyricularia, fueron repicadas sobre trozos de papel filtro colocados en placas de Petri con medio PDA, cultivadas por 8 días a 25°C; posteriormente los trozos de papel fueron retirados y colocados en placas de Petri estériles para el secado y posterior conservación a -18°C (Aricapa y Correa 1994).
Inoculación de materiales de trigo
Para la prueba de patogenicidad, se seleccionaron 9 aislados, cada uno de un hos- pedero diferente, más un aislado de trigo como testigo. En la tabla 2, se presenta la lista de aislados seleccionados. Estos aislados fueron repicados a placas de Petri con medio de cultivo Agar-Harina de avena, cultivados por 12 días a 25°C con foto- periodo de 12 horas, posteriormente las colonias fueron aplastadas con una varilla de vidrio, y las placas colocadas bajo luz fluorescente constante por 48 horas para favorecer la esporulación (Marangoni et al. 2013). Para preparar la suspensión de conidios, se agregó 10 ml de agua destilada estéril a cada placa y las colonias fueron raspadas con ayuda de un cepillo, luego se ajustó la concentración a 5.104 conidios/ ml-1 con un hemacitómetro.
Las variedades de trigo inoculadas fueron Canindé 1 (Resistente), Canindé 11(Susceptible) e Itapúa 70 (Moderadamente susceptible). La inoculación se realizó asperjando las espigas con las suspensiones de cada aislado.
Tabla 2. Aislados seleccionados para las pruebas de patogenicidad con su código y origen. Caacupé, Paraguay, 2016.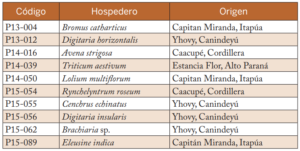
Para cada aislado se inocularon 4 macetas con tres plantas, de cada variedad. Pos- terior a la inoculación las plantas se mantuvieron en una cámara climatizada con 27°C de temperatura y 80% de humedad (Chávez et al. 2017). La evaluación se realizó 15 días después de la inoculación indicando la reacción de compatibilidad (+) e icompatibilidad (-).
RESULTADOS
De las muestras colectadas en el departamento de Itapúa, se constató la presencia de Pyricularia en hojas de Bromus catharticus, Cenchrus echinatus, Chloris gayana, Digita- ria horizontalis y Eleusine indica, así como en hojas e inflorescencias de Avena strigosa y Lolium multiflorum; y en inflorescencias de Cyperus diffusus. En el departamento de Alto Paraná, son hospederos C. echinatus y D. horizontalis; y en el departamento de Canindeyú, A. strigosa, Brachiaria sp., C. echinatus, D. insularis y D. horizontalis. En el departamento de Cordillera, se constató la presencia del hongo en las tres especies muestreadas, A. strigosa, Brachiaria sp. y Rynchelyntrum roseum.
Tabla 3. Compatibilidad de la infección de los diferentes aislados en las tres variedades inoculadas. Caacupé, 2016.
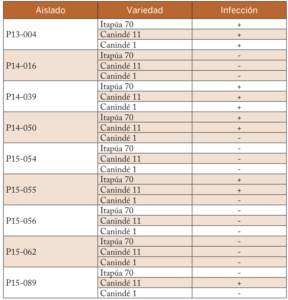
De los nueve aislados seleccionados, los aislados de Cenchrus echinatus, Eleusine indica, Lolium multiflorum y Bromus catharticus causaron infección en las varieda- des inoculadas. La compatibilidad de las infecciones de los diferentes aislados se presentan en la tabla 3.
CONCLUSIÓN
Se constató la presencia de Pyricularia en las siguientes especies: Avena strigosa, Brachiaria sp., Bromus catharticus, Lolium multiflorum, Chloris gayana, Cyperus di- ffusus, Cenchrus echinatus, Eleusine indica, Digitaria insularis, Digitaria horizontalis y Rynchelyntrum roseum. Si bien en muchas de las especies colectadas, no se observó la presencia del hongo, no se las debe descartar como posibles hospederos alternativos, ya que los numerosos reportes demuestran que Pyricularia posee un amplio rango de hospedantes, y se debe seguir estudiando los mismos.
Los aislados de Cenchrus echinatus, Lolium multiflorum, Bromus catharticus y Eleusine indica causan infección al trigo, teniendo en cuenta que solo se estudió un aislado de cada hospedero, es necesario ampliar el número de aislados estudiados.
REFERENCIAS BIBLIOGRÁFICAS
_ Barnett, HL; Hunter, BB. 1998. Ilustrated genera imperfect fungi. 4 ed. The American Phytopathological society. Minnesota, US. 218 p.
_ Chávez, A; Cazal, C; Kohli, MM. 2017. Diferencia en la reacción a Pyricularia oryzae de materiales de trigo en los estadios vegetativo y reproductivo. Investigación Agraria 19(1):56-63.
_ Gutiérrez, S; Mazzanti de Castañon, M; Galmarini, M. 2000. Avances en el conoci- miento de hospederos espontáneos de Magnaporthe oryzae en Argentina. (en línea). Co- municaciones científicas y tecnológicas, Universidad Nacional del Nordeste. Consultado 06 ago. 2013. Disponible en: www.unne.edu.ar/unnevieja/Web/cyt/cyt/2000/5…/a_pdf/a_043.pd
_ Lima, MIPM. 2004. Giberela ou Brusone? Orientaçoes para a identificação correta des- sas enfermidades em trigo e em cevada. (en línea). Documentos on line. Embrapa Trigo. Consultado 30 abr. 2013. Disponible en: http://www.cnpt.embrapa.br/biblio/do/p_do40. htm
_ Marangoni, M; Nunes, M; Fonseca, N; Metha, Y. 2013. Pyricularia blast on White oats – a new threat to wheat cultivation. Tropical Plant Pathology. 38(3):198-202.
_ Mehta, YR; Nunes, MP; Oliveira, JC. 2006. Occurrência de brusone em aveia no Estado do Paraná. In: Resultados Experimentais. XXVI Reunião da Comissão Brasileira de Pes- quisa de Aveia, 4–6 de abril, FAPA, Guarapuava, Paraná, 55–57.
_ Prabhu, AS; Filippi, MC; Castro, N. 1992. Pathogenic variation among isolates of Mag- naporthe oryzae infecting rice, wheat, and grasses in Brazil. Tropical Pest Management. 38: 367-371.
_ Reis, EM; Casa, RT; Forcelini, CA. 2005. Doenças do trigo (Triticum aestivum L.). In Kimati, H; Amorim, L; Rezende, J; Bergamin, A; Camargo, L. Manual de Fitopatologia: doenças das plantas cultivadas. 4 ed. São Paulo, BR. Ceres. v. 2, 663 p.
_ Tagle, AG; Chuma I; Tosa, Y. 2014. Rmg7, a new gene for resistance to Triticum isolates of Pyricularia oryzae identified in tetraploid wheat. Genetics and resistance. V 105(4) 495-499.
_ Urashima, AS; Igarashi, S; Kato, H. 1993. Host range, mating type, and fertility of Mag- naporthe oryzae from wheat in Brazil. Plant Disease, 12: 11–16.
_ Valent, B; Cruz, CD; Peterson, G; Bockus, WW; Farman, M; Pedley, K; Whitley, R; Navia-Urrutia, M; Trick, HN; Maciel, JLN; Oliveira-Garcia, E; Stack, J. 2016. Wheat blast: Biology, genetics and genomics. In: Madeiros Del Ponte, E; Bergstrom, G; Pavan, W; Lazzaretti, A; Cunha Fernandes, JM. Book of Abstracts. 5th International Symposium on Fusarium head blight. 2nd International Workshop on Wheat Blast. Universidade de Paso Fondo, RS. BR.
_ Verzignassi, JR; Poltronieri, LS; Benchimol, RL; Santos de França, SK; de Arruda, Eu- des; Dornelas, C. 2012. Magnaporthe oryzae: novo patógeno em Brachiaria brizantha cv. Marandú no Pará. Summa Phytopathologica 38 (3): 254.
_ Viedma, LQ; Morel, W. 2002. Añublo o Pyricularia del Trigo. Díptico. MAG/DIA/ CRIA. Programa de Investigación de Trigo, CRIA, Capitán Miranda, Itapúa.